

Спикер: Андрей Юрченко, кандидат биологических наук, Institut Gustave Roussy (Франция), Институт цитологии и генетики СО РАН.
Чтобы принять участие в практической международной конференции KSIDAY 2022, которая в этом году пройдёт 2 ноября, подавайте заявку на подключение к онлайн-трансляции мероприятия по ссылке: https://ksiday.ru/.
Хотел бы рассказать о последних исследованиях в области популяционной генетики. Работа выполнялась в основном на базе Института ЦиГ РАН, а также вместе с коллегами из других институтов.
Напомню об основных генетических линиях и путях доместикации КРС. Общепринятая теория гласит о том, что около 500 лет назад предковые евразийские популяции тура, предка всех коров, разделились на европейскую и азиатскую части. Затем около 8 500 лет назад из азиатской части была проведена доместикация части животных. Из них в итоге получились азиатские индициновые породы скота, которые мало представлены в России, но в Индии и Азии они играют огромную роль.
Примерно в то же время около 10 000 лет назад в районе Ближнего Востока был доместицирован европейский тур, который дал начало основным породам европейского тауринового скота. К нему относится большинство европейских пород, российские породы в том числе.
Также существует линия — африканский тауриновый скот. Эта линия отделилась от европейских тауринов около 6 000 лет назад и затем развивалась в Африке. Потом около 700 лет назад эти африканские тауриновые породы стали скрещиваться с завезёнными индициновыми породами. Сейчас в Африке имеется большое разнообразие скота.
Для нас наибольший интерес представляют пока европейские тауриновые породы. Наша работа начиналась с того, что три года назад мы собрали 297 животных, которые принадлежали к 18 породам, основным породам российского скота. Генотипировали их на основе 40 000 «снипов» на панели, а затем сравнили эти породы с огромной коллекцией из 2 000 животных и 130 пород мирового генофонда, которые были опубликованы до нас.
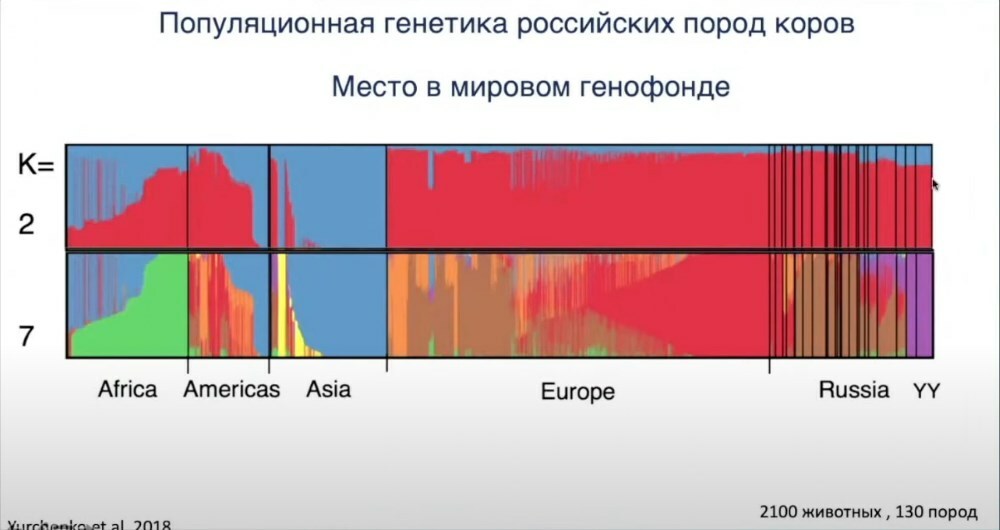
Вы видите, что, если мы производим кластеризацию особей на основе генотипов, мы можем тестировать, относятся ли те или иные животные к тем или иным породам, группам. Например, задавая количество кластеров, мы можем задавать вопрос: если мы разделим всех этих животных на два кластера, то какие основные кластеры генетически не распадутся? Здесь мы видим, что они распадаются в основном в соответствии с географией. Европа — красный кластер, Азия — синий, в Африке много гибридов. Если мы попробуем протестировать наличие семи кластеров, то увидим, что азиатские породы представлены индициновыми породами, российские породы скота окрашены в коричневый, фиолетовый или красный. Эти же красные и коричневые кластеры мы наблюдаем в Европе. Но, как видите, фиолетового нет нигде, кроме России, — это породы, относящиеся к другой ветви: татаро-монгольский скот, якутская и бурятская породы.
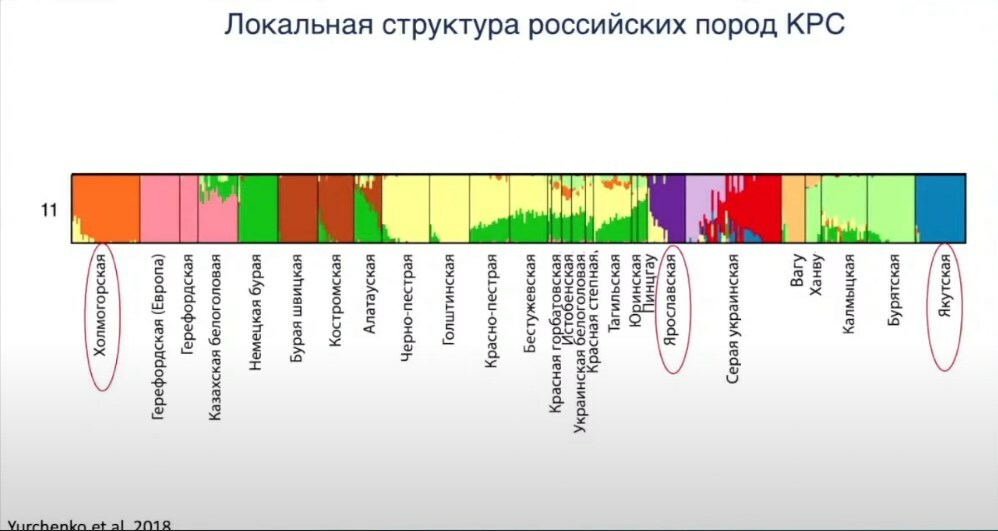
Также мы интересовались таким вопросом, как локальность таких пород КРС. Особенно было интересно найти какие-то породы, которые уникальны в России и не представлены больше нигде в мире. Мы знаем, что на протяжении XVIII—XX веков огромное количество пород ввозилось на территорию России и Советского Союза и активно смешивалось с нашими древними породами. Например, если мы посмотрим на генетический состав костромской породы, то увидим, что она окрашена таким же цветом, как бурая швицкая, то есть они очень близки. Естественно, костромская порода не вымерла, но если её сравнить с европейской породой бурой швицкой, они крайне близки генетически, потому что проводились частые скрещивания на улучшение породы.
Такая же история с казахской белоголовой породой. Она смешана с герефордской породой и генетически близка к ней. Красно-пёстрая порода очень похожа на голштинскую. Вообще этот большой кластер пород — бестужевская, красно-горбатовская, чёрно-пёстрая, — они очень сильно подвержены влиянию голштинов, которые раньше специально завозились для улучшения российских пород.
Нам удалось найти две породы из тауриновых — холмогорская и ярославская, — которые не были подвержены влиянию завезённых европейских пород и развивались, по всей видимости, со времён раннего Средневековья где-то на Севере России. Они представляют собой остатки того уникального генофонда, который когда-то присутствовал.
Якутская окрашена совсем другим цветом, она не кластеризуется ни с одной из этих пород. Бурятская российская порода скота считалась вымершей до недавнего времени, но было найдено её стадо в Монголии. Затем завезены в Россию и разводятся. Бурятская группируется рядом с калмыцкой и ханву.
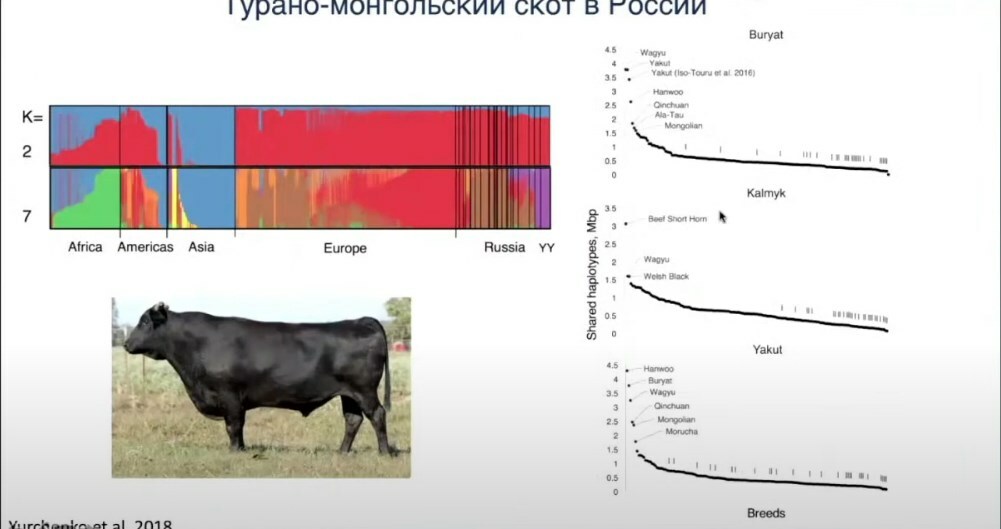
Если говорить о бурятской, калмыцкой и якутской породах, которые мы исследовали в нашей первой работе. Чем они уникальны и почему не кластеризуются с обычными европейскими тауриновыми породами? Я проводил анализ гаплотипов. Если какие-то животные более-менее близки с другими породами, то по анализу гаплотипов, сходных участков генома мы можем это найти. Если мы берём якутскую породу скота, то наибольшее количество общих гаплотипов она имеет с такими породами, как ханву, бурятская, вагу и другими азиатскими. По бурятской мы наблюдаем то же самое — она ближе всего к ханву, вагу.
Увидев эти интересные вещи, мы решили более плотно провести анализ татаро-монгольских пород. Для этого в нашей недавней работе мы собрали генотипы около 2 700 животных, которые представляли собой 198 пород, из них 513 животных представляли 23 турано-монгольские породы. Турано-монгольские породы — это интересное направление, многие китайские учёные считают, что эти коровы были доместицированы отдельным этапом где-то 10 000 лет назад в районе Северного Китая, затем они там распространились. Далеко не все учёные поддерживают эту версию.

Если мы построим филогенетическое дерево всех известных пород и окрасим в разный цвет, то увидим, что турано-монгольский скот группируется в корне этого дерева. Он действительно находится где-то отдельно от европейских тауринов.

Если мы проводим генетическую кластеризацию, то ситуация получается ещё более интересной. Если мы кластеризуем всё наше разнообразие только на две группы, то оно разделяется только на индициновый и тауриновый скот, синее и красное. Если идём дальше и разделяем, у нас добавляется африканский и тауриновый скот. И если разделяем на четыре части, то вот эта большая фиолетовая четвёртая часть описывается разнообразием турано-монгольского скота. То есть это породы из России — якутская, бурятская, из Китая, — целый набор пород, в том числе тибетские. Они действительно имеют под собой мощный генетический компонент, который делает их уникальными и выделяет из многих других пород скота.
Печально, что, как видите, фиолетового кластера, полностью фиолетовых животных у нас практически здесь нет. Все остальные имеют ту или иную степень смешения с европейскими тауринами или индициновым скотом. Это подтверждают записи, известные наблюдения, что в XX веке большая часть турано-монгольских коров была случайно смешана с какими-то европейскими или азиатскими породами.
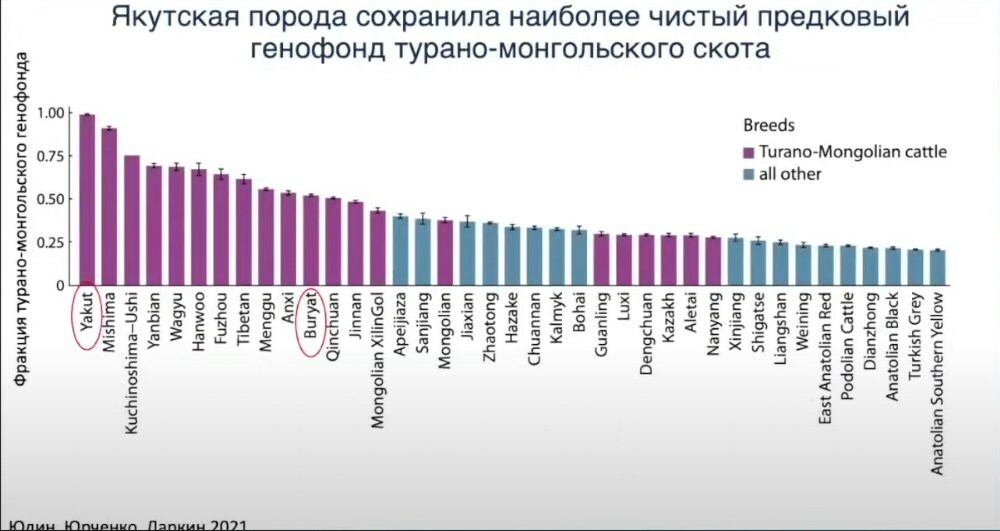
Мы решили оценить чистоту турано-монгольского скота. То есть если мы строим модель, допустим, где мы можем оценить фракцию турано-монгольского генофонда в той или иной породе, то получается интересный результат. Вот здесь сортированы по фракциям турано-монгольского генофонда от совсем «чистых» до совсем «грязных». Турано-монгольские показаны только фиолетовым, синие — имеют примесь турано-монгольских, но ими не являются.
Самое интересное, что единственной по-настоящему чистой породой является якутская порода скота, которая разводится сейчас только в России. Она представляет собой реликт когда-то важного азиатского турано-монгольского генофонда. Следом за ней идёт японская порода мишима, которая разводится на каком-то острове, она на 90% представлена фракцией турано-монгольского генофонда. Бурятская порода скота на 60% определяется этим генофондом, она успела где-то «нахватать» генов от других пород и генетических линий.
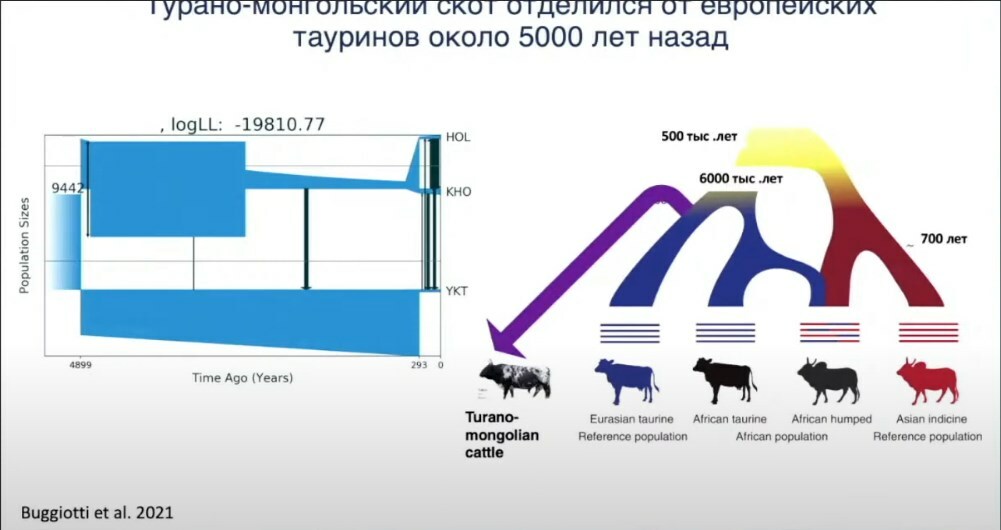
Мы секвенировали полные геномы турано-монгольского скота, представленного в основном якутской ветвью. Оказалось, что если мы вскроем модель на основе геномов по времени разделения, то можно сказать, что турано-монгольская, по крайней мере якутская ветвь, отделилась от остального евразийского тауринового скота около 5 000 лет назад. Если мы это наложим на рисунок, который я уже показывал, то у нас вместо двух тауриновых древних ветвей — евразийской и африканской — добавляется ещё и турано-монгольская ветвь, которая отделилась чуть позже африканской и изолированно развивалась в районе Азии, Китая, современной Республики Якутия. Что немного печально, несмотря на то, что якутская корова оказалась настолько уникальной.
Если мы посмотрим такую метрику, как среднее число регионов гомозиготности на животное и среднюю длину регионов гомозиготности на животное, якутская корова среди российских пород, к сожалению, лидирует. Она является достаточно сильно инбредной. Видимо, это связано с событиями Великой Отечественной войны, когда огромное количество якутского скота было отправлено на мясо. Сейчас их пытаются снова развивать. Думаю, генетическая селекция здесь будет как нельзя кстати. Если отбирать животных, разнообразных по генотипу, и скрещивать, то, думаю, уровень якутской крайней гомозиготности можно со временем опустить и повысить разнообразие этих животных.
Мы также занимаемся поиском следов отбора в геномах различных российских пород скота. Например, чем уникальны якутская и бурятская породы? Якутская порода — единственная порода, которая живёт за полярным кругом, она легко выдерживает морозы до -50 градусов. Это крайне устойчивое животное, не требовательное к кормам. Естественно, у неё должны быть адаптации, которые позволили ей добиться такой невероятной устойчивости. Бурятская корова тоже из той же линии турано-монгольского скота. Если сравнивать регионы, которые находятся под отбором и в той, и другой породах, мы увидим, что гены разные. На самом деле каждая порода уникальна, каждая из них имеет свои отобранные генетические варианты, которые являются уникальной частью генофонда, который нужно сохранять, изучать и понимать. К сожалению, для многих этих генов мы не знаем точных функций и какие варианты вызывают адаптации к тем или иным условиям. Но эта работа сейчас проводится и является важной.
На примере сибирских герефордов расскажу ещё об одной нашей работе. Сибирские герефорды давно живут в условиях сибирского климата и успели адаптироваться. Мы проводили работу, где взяли 197 животных, зимой на пике морозов у них на протяжении нескольких дней измерялась температура тела. Затем мы разделили животных на группы, которые хорошо переносят мороз и которые переносят его хуже — у них скачет температура, им труднее адаптироваться к морозу. Генотипировали их и попытались найти регионы генома, которые могли бы объяснить устойчивость к низким температурам. Был найден один из регионов генома на основе гаплотипов, на основе поиска ассоциаций, там находилось два гена — MSANTD4 и GRIA4. Затем я посмотрел, не находятся ли эти регионы под отбором. Важно понимать, что мутации, которые там находятся, они не фиксированы в этой популяции. Часть их имеет, часть не имеет. Но тем не менее в этом регионе было заметно, что отбор либо проходил в какие-то суровые зимы, либо до сих пор ещё идёт, и животные постепенно отбираются.
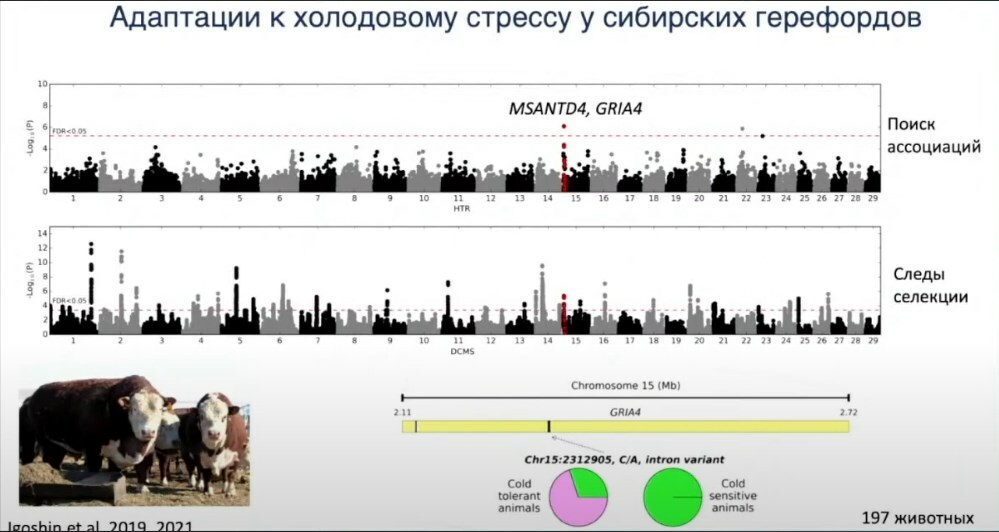
Потом мы секвенировали ещё 12 животных, которые показывали собой контрастирующие фенотипы, устойчивые и неустойчивые к холоду, грубо говоря. Секвенировали полные геномы и нашли мутацию, которая наиболее сильно ассоциирована с устойчивостью к холоду — мутация в гене GRIA4. Здесь видно, что у животных, которые были чувствительны к холоду, в основном присутствует аллель A, а у нечувствительных — аллель С.
Такие примеры могут использоваться для геномной селекции — генотипирование будет проводиться намного проще. Достаточно посмотреть на отдельные замены, а не секвенировать полный геном.
Недавнее исследование. Мы секвенировали геном якутской коровы и обнаружили, что в одном из генов, NRAP, у неё с высокой частотой присутствует специфический гаплотип. В других породах, таких как холмогорская и голштинская, этого гаплотипа почти нет. Этот гаплотип всегда находился с одной мутацией — она была крайне необычна тем, что была с высокой частотой у якутской коровы. Среди 5 000 проверенных геномов из проекта генома быков мы больше ни у кого не нашли эту мутацию. Также если сравнить с геномом других животных, таких как человек, мыши, свинья и медведь, то эта мутация крайне редка, её почти нет. Потом мы поискали информацию, провели эволюционное сравнение, и оказалось, что эта мутация в этом гене находится у животных, которые впадают в спячку. Спячка — это очень сложный процесс, он относится не только ко сну, это прежде всего сокращение активности метаболизма, специальное понижение температуры тела и снижение нагрузки на сердце. По всей видимости, эта уникальная мутация каким-то образом помогает якутской корове адаптироваться к холоду. Сейчас дополнительно проводится молекулярное исследование, чтобы лучше понять эту функцию. Ген этот в основном работает в сердце, и у людей, которых есть «поломки» в этом гене, возникают такие проблемы, как аритмия и проблемы с кровяным давлением.
Заключение.
Несмотря на долгую историю гибридизации с европейскими породами, российские обладают специфическими гаплотипами, приобретёнными в процессе селекции на территории России. Даже если породы достаточно сильно смешаны с европейскими породами, тем не менее в любом случае очень во многих породах имеются уникальные гаплотипы, которые уже были отобраны на территории России.
Холмогорская и ярославская породы являются генетически уникальными породами российского происхождения, они должны охраняться.
Якутская порода представляет собой наиболее чистый пример азиатского турано-монгольского скота, который был подвержен сильному влиянию коммерческих пород в XX веке и, по сути, является реликтом древнеазиатского скота.
Следы адаптации к холодному климату прослеживаются у многих российских пород и могут быть полезны для улучшения коммерческих пород на территории России.


